Типы мышечных сокращений. Виды и режимы мышечного сокращения Снижение мышечных усилий называется
Типы мышечных сокращений. По способу укорочения мышц различают три типа мышечных сокращений:
1) изотоническое , при котором волокна укорачиваются при постоянной внешней нагрузке, в реальных движениях проявляется редко (так как мышцы укорачиваясь вместе с тем меняют своё напряжение);
2) изометрическое – это тип активации, при котором мышца развивает напряжение без изменения своей длины . На нём построена так называемая статическая работа двигательного аппарата человека. Например, в режиме изометрического сокращения работают мышцы человека, который подтянулся на перекладине и удерживает своё тело в этом положении;
3) ауксотоническое или анизотоническое – это режим, при котором мышца развивает напряжение и укорачивается . Именно этот тип мышечных сокращений обеспечивает выполнение двигательных действий человека.
У анизотонического сокращения две разновидности сокращения мышцы: в преодолевающем и уступающем режимах.
В преодолевающем режиме мышца укорачивается в результате сокращения (например, икроножная мышца бегуна укорачивается в фазе отталкивания).
В уступающем режиме мышца растягивается внешней силой (например, икроножная мышца спринтера при взаимодействии ноги с опорой в фазе амортизации).
На рисунке 1 изображена динамика работы мышцы в преодолевающем и уступающем режимах.
Правая часть кривой отображает закономерности преодолевающей работы, при которой возрастание скорости сокращения мышцы вызывает уменьшение силы тяги.
В уступающем режиме наблюдается обратная картина: увеличение скорости растяжения мышцы сопровождается увеличением силы тяги (что является причиной многочисленных травм у спортсменов, например, разрыв ахиллова).
При скорости, равной нулю, мышцы работают в изометрическом режиме.
Для движения звена в суставе под действием мышечных сил важны не сами силы, а создаваемые ими моменты сил , поскольку движение звена – это ни что иное, как вращение относительно оси, проходящей через сустав. Поэтому разновидности работы мышц можно выразить в терминах моментов сил: если отношение момента внутренних сил к моменту внешних рано единице, режим сокращения будет изометрическим, если больше единицы – преодолевающим, если меньше единицы – уступающим. Поддержку сустава можно обеспечить спортивным тейпом .
Групповое взаимодействие мышц. Существует два вида группового взаимодействия мышц: синергизм и антагонизм.
Мышцы-синергисты перемещают звенья тела в одном направлении. Например, в сгибании руки в локтевом суставе участвуют двуглавая мышца плеча, плечевая и плечелучевая мышцы. В результате синергического взаимодействия мышц увеличивается результирующая сила действия.
Мышцы-антагонисты имеют разнонаправленное действие: если одна из них выполняет преодолевающую работу, то другая – уступающую. Мышцы обеспечивают возвратно-вращательные движения звеньев тела, поскольку каждая из них работает только на сокращение; высокую точность двигательных действий, так как звено необходимо не только привести в движение, но и затормозить в нужный момент. Антагонисты состоят из пары: агонист (сгибатель) – антагонист (разгибатель).
Мощность и эффективность мышечного сокращения. По мере увеличения скорости мышечного сокращения сила тяги мышцы, функционирующей в преодолевающем режиме, снижается по гиперболическому закону (см. рис. 1). Известно, что механическая мощность равна произведению силы на скорость (N = F V). Существует сила и скорость, при которых мощность мышечного сокращения наибольшая; этот режим возникает, когда и сила, и скорость составляют примерно 30 % от максимально возможных величин.
Накопление энергии упругой деформации в растянутых мышцах и сухожилиях. Когда сокращению мышц предшествует фаза растяжения, производимые силы, мощность и работа достигают больших величин по сравнению с сокращением без предварительного растяжения. После растяжения скорость сокращения увеличивается за счёт скорости восстановления упругих компонентов мышцы.
Растяжение мышечно-сухожильной системы позволяет также накапливать и использовать энергию упругой деформации. Было подсчитано, что ахиллово сухожилие растягивается на 18 мм во время бега со средней скоростью , при этом накапливается энергия в 42 Дж. Нелинейная зависимость между величиной растяжения и накапливаемой энергией показывает, что при больших растяжениях накапливается больше энергии, чем при малых. Эластичное растяжение внесёт значительный вклад в мышечную деятельность, только если за активным мышечным растяжением немедленно последует преодолевающий режим сокращения мышцы. Более высокая результативность прыжка с подседом по отношению к прыжку из статической позы показывает преимущество предварительного растяжения мышц.
Скелетная мускулатура является составной частью опорно-двигательного аппарата человека. При этом мышцы выполняют следующие функции:
1) обеспечивают определенную позу тела человека;
2) перемещают тело в пространстве;
3) перемещают отдельные части тела относительно друг друга;
4) являются источником тепла, выполняя терморегуляционную функцию.
Свойства скелетной мышцы :
1) Возбудимость - способность отвечать на действие раздражителя изменением ионной проводимости и мембранного потенциала. В естественных условиях этим раздражителем является медиатор ацетилхолин, который выделяется в пресинаптических окончаниях аксонов мотонейронов. В лабораторных условиях часто используют электрическую стимуляцию мышцы. При электрической стимуляции мышцы первоначально возбуждаются нервные волокна, которые выделяют ацетилхолин, т.е. в данном случае наблюдается непрямое раздражение мышцы. Это обусловлено тем, что возбудимость нервных волокон выше мышечных. Для прямого раздражения мышцы необходимо применять миорелаксанты - вещества, блокирующие передачу нервного импульса через нервно-мышечный синапс;
2) Низкая проводимость (10-13 м/с) - способность проводить потенциал действия вдоль и вглубь мышечного волокна по Т-системе;
3) Сократимость - способность укорачиваться или развивать напряжение при возбуждении;
4) Эластичность - способность развивать напряжение при растягивании.
5) Рефрактерность – отсутствие или снижение возбудимости нерва или мышцы после предшествующего возбуждения. Занимает по времени больший отрезок, чем у нервного волокна.
6) Лабильность – функциональная подвижность, скорость протекания элементарных циклов возбуждения в нервной и мышечной тканях
Основными показателями, характеризующими деятельность мышц, являются их сила и работоспособность.
Сила мышц . Сила - мера механического воздействия на мышцу со стороны других тел, которая выражается в ньютонах или кг-силах. При изотоническом сокращении в эксперименте сила определяется массой максимального груза, который мышца может поднять (динамическая сила ); при изометрическом - максимальным напряжением, которое она может развить (статическая сила).
Одиночное мышечное волокно развивает напряжение в 100-200 кг-сил во время сокращения.
Степень укорочения мышцы при сокращении зависит от силы раздражителя, морфологических свойств и физиологического состояния. Длинные мышцы сокращаются на большую величину, чем короткие.
Незначительное растяжение мышцы, когда напрягаются упругие компоненты, является дополнительным раздражителем, увеличивает сокращение мышцы, а при сильном растяжении сила сокращения мышцы уменьшается.
Напряжение, которое могут развивать миофибриллы, определяется числом поперечных мостиков миозиновых нитей, взаимодействующих с нитями актина, так как мостики служат местом взаимодействия и развития усилия между двумя типами нитей. В состоянии покоя довольно значительная часть поперечных мостиков взаимодействует с актиновыми нитями. При сильном растяжении мышцы актиновые и миозиновые нити почти перестают перекрываться и между ними образуются незначительные поперечные связи.
Величина сокращения снижается также при утомлении мышцы.
Изометрически сокращающаяся мышца развивает максимально возможное для нее напряжение в результате активации всех мышечных волокон. Такое напряжение мышцы называют максимальной силой . Максимальная сила мышцы зависит от числа мышечных волокон, составляющих мышцу, и их толщины. Они формируют анатомический поперечник мышцы, который определяется как площадь поперечного разреза мышцы, проведенного перпендикулярно ее длине. Отношение максимальной силы мышцы к ее анатомическому поперечнику называется относительной силой мышцы, измеряемой в кг/см2.
Физиологический поперечник мышцы - длина поперечного разреза мышцы, перпендикулярного ходу ее волокон.
В мышцах с параллельным ходом волокон физиологический поперечник совпадает с анатомическим. У мышц с косыми волокнами он будет больше анатомического. Поэтому сила мышц с косыми волокнами всегда больше, чем мышц той же толщины, но с продольными волокнами. Большинство мышц домашних животных и особенно птиц с косыми волокнами перистого строения. Такие мышцы имеют больший физиологический поперечник и обладают большей силой.
Наиболее сильными являются многоперистые мышцы, затем идут одноперистые, двухперистые, полуперистые, веретенообразные и продольноволокнистые.
Много, -одно, -и двухперистые мышцы имеют большую силу и выносливость (мало утомляются), но ограниченную способность к укорачиванию, а остальные виды мышц хорошо укорачиваются, но быстро утомляются.
Сравнительным показателем силы разных мышц является абсолютная мышечная сила - отношение максимальной силы мышцы к ее физиологическому поперечнику, т.е. максимальный груз, который поднимает мышца, деленный на суммарную площадь всех мышечных волокон. Она определяется при тетаническом раздражении и при оптимальном исходном растяжении мышцы. У сельскохозяйственных животных абсолютная сила скелетных мышц колеблется от 5 до 15 кг-сил, в среднем 6-8 кг-сил на 1см2 площади физиологического поперечника. В процессе мышечной работы поперечник мышцы увеличивается и, следовательно, возрастает сила данной мышцы.
Работа мышц . Работа мышц внешне выражается либо в фиксации части тела, либо в движении. В первом случае говорят о так называемой статической работе, а во втором – о динамической работе.
Статическая работа мышц есть следствие равенства моментов сил и называется еще удерживающей работой. При такой работе форма мышцы, ее размеры, возбуждение и напряжение относительно постоянны.
Динамическая работа мышц сопровождается движением и есть следствие разности моментов сил. В зависимости от того, какой момент окажется большим, различают два вида динамической работы мышц: преодолевающую и уступающую. Превалирование момента силы мышцы или группы мышц приводит к преодолевающей работе, а уменьшение момента силы мышцы – к уступающей работе.
Различают еще баллистическую работу мышц, которая является разновидностью преодолевающей работы: мышца совершает быстрое сокращение и последующее расслабление, после которого костное звено продолжает движение по инерции.
(10) Виды и режимы сокращения скелетной мышцы. Одиночное мышечное сокращение, его фазы. Тетанус и его виды. Оптимум и пессимум раздражения.
Виды сокращений .
У скелетной мышцы выделяют одиночное сокращение и суммированное сокращение (тетанус).
Одиночное сокращение - это сокращение, которое возникает на одиночный стимул, достаточный для вызова возбуждения мышцы.
Фазы одиночного мышечного сокращения :
Латентный период. Представляет собой сумму временных задержек, обусловленных возбуждением мембраны мышечного волокна, распространением ПД по Т-системе внутрь волокна, образованием инозитолтрифосфата, повышением концентрации внутриклеточного кальция и активации поперечных мостиков. Для портняжной мышцы лягушки латентный период составляет около 2 мс.
Период укорочения, или развития напряжения.
Период расслабления, когда уменьшается концентрация ионов Са2+ и головки миозина отсоединяются от актиновых филаментов.
При воздействии на мышцу ритмических раздражений высокой частоты наступает сильное и длительное сокращение мышцы, которое называется тетаническим сокращением , или тетанусом.
Тетанус может быть зубчатым (при частоте раздражений 20-40 Гц) или гладким (при частоте 50 Гц и выше). Амплитуда тетанического сокращения в 2–4 раза выше амплитуды одиночного сокращения при той же силе раздражения.
Гладкий тетанус возникает тогда, когда очередной импульс раздражения действует на мышцу до начала фазы расслабления. При очень большой частоте раздражений каждое очередное раздражение будет попадать на фазу абсолютной рефрактерности и мышца вообще не будет сокращаться. Высота мышечного сокращения при тетанусе зависит от ритма раздражения, а также от возбудимости и лабильности, которые изменяются в процессе сокращения мышцы. Тетанус наиболее высокий при оптимальном ритме, когда каждый последующий импульс действует на мышцу в фазу экзальтации, вызванной предыдущим импульсом. В этом случае создаются наилучшие условия (оптимум силы и частоты раздражения, оптимум ритма) для работы мышцы.
При тетанических сокращениях мышечные волокна утомляются больше, чем при одиночных сокращениях. Поэтому даже в пределах одной мышцы происходит периодическая смена частоты импульсации (вплоть до полного исчезновения) в разных двигательных единицах.
Для скелетной мышцы характерны два основных режима сокращения - изометрический и изотонический.
Изометрический режим проявляется в том, что в мышце во время ее активности нарастает напряжение (генерируется сила), но из-за того, что оба конца мышцы фиксированы (например, мышца пытается поднять большой груз), она не укорачивается.
При изотоническом режиме мышца первоначально развивает напряжение (силу), способную поднять данный груз, а потом укорачивается (меняет свою длину, сохраняя напряжение, равное весу поднимаемого груза).
Оптимум – уровень силы или частоты раздражений, при котором осуществляется максимальная деятельность органа или ткани. Явление О. описано Н. Введенским, который на нервно-мышечном препарате лягушки установил, что нарастание до некоторого предела частоты или силы раздражений усиливает длительное, слитное сокращение мышцы - тетанус. О. объясняют тем, что в этих случаях каждое последующее раздражение падает на мышцу в период повышенной её возбудимости, вызванной предыдущим раздражением.
Пессимум - угнетение деятельности органа или ткани, вызываемое чрезмерной частотой или силой наносимых раздражений. Это явление было описано Н. Введенским. Исследуя особенности проведения нервного импульса в нервно-мышечном препарате лягушки, он обнаружил, что усиление тетануса, вызываемое постепенным возрастанием частоты или силы раздражений, при дальнейшем их учащении или усилении, внезапно сменяется расслаблением мышцы и полным торможением её активности. Введенский трактовал это явление с позиций разработанной им теории парабиоза . Согласно этой теории, работоспособность нервных окончаний, передающих импульсы мышце, после прохождения волны возбуждения резко падает, и для восстановления их работоспособности требуется некоторое время (в нервно-мышечном препарате икроножной мышцы лягушки - 0,02-0,03 сек). Это время определяет функциональные возможности нервных окончаний - их лабильность . Если интервал между раздражениями меньше этого необходимого периода, то есть если он превышает лабильность нервных окончаний, в них развивается своеобразное стойкое нераспространяющееся возбуждение - парабиоз , блокирующее проведение нервных импульсов к мышце и тормозящее тем самым её активность, предохраняя от переутомления. Описываемое явление носит обратимый характер: снижение интенсивности раздражения восстанавливает мышечное сокращение.
В зависимости от характера сокращений мышцы различают три их вида: изометрическое, изотоническое и ауксотоническое .
Ауксотоническое сокращение мышцы заключается в одновременном изменении длины и напряжения мышцы. Этот вид сокращения характерно для натуральных двигательных актов и бывает двух видов: эксцентрическое, когда напряжение мышцы сопровождается ее удлинением - например, в процессе приседания (опускания), и концентрическое, когда напряжение мышцы сопровождается ее укорочением - например, при разгибании нижних конечностей после приседания (подъем).
Изометрическое сокращение мышцы - когда напряжение мышцы возрастает, а длина ее не изменяется. Этот вид сокращения можно наблюдать в эксперименте, когда оба конца мышцы зафиксированы и отсутствует возможность их сближения, и в естественных условиях - например, в процессе приседания и фиксации положения.
Изотоническое сокращение мышцы заключается в укорочении мышцы при ее постоянном напряжении. Этот вид сокращения возникает, когда сокращается ненагруженная мышца с одним закрепленным сухожилием, не поднимая (не перемещая) никакого внешнего груза либо поднимая груз без ускорения.
В зависимости от длительности сокращений мышцы выделяют два их вида: одиночное и тетаническое.
Одиночное сокращение мышцы возникает при однократном раздражении нерва или самой мышцы. Обычно мышца укорачивается на 5-10% от исходной длины. На кривой одиночного сокращения выделяют три основных периода: 1) латентный - время от момента нанесения раздражения до начала сокращения; 2) период укорочения (или развития напряжения) ; 3) период расслабления . Продолжительность одиночных сокращений мышц человека вариабельна. Например, у камбаловидной мышцы она составляет 0,1 с. В латентный период возникает возбуждение мышечных волокон и его проведение вдоль мембраны. Соотношения длительности одиночного сокращения мышечного волокна, его возбуждения и фазовые изменения возбудимости мышечного волокна показаны на рис. 24.
Длительность сокращения мышечного волокна значительно дольше таковой ПД потому, что необходимо время на работу Са-насосов для возвращения Са 2+ в СПР и окружающую среду и большей инерционности механических процессов по сравнению с электрофизиологическими.
Рис. 24. Соотношение времени возникновения ПД (А) и одиночного сокращения (Б) медленного волокна скелетной мышцы теплокровного. Стрелка – момент нанесения раздражения. Время сокращения быстрых волокон в несколько раз короче
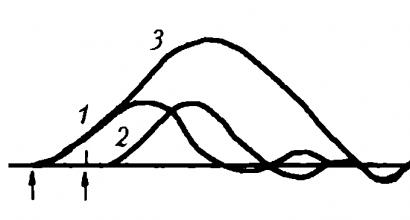
Тетаническое сокращение - это длительное сокращение мышцы, возникающее под действием ритмического раздражения, когда каждое последующее раздражение или нервные импульсы поступают к мышце, пока она еще не расслабилась. В основе тетанического сокращения лежит явление суммации одиночных мышечных сокращений (рис. 25) - увеличение амплитуды и длительности сокращения при нанесении на мышечное волокно или целую мышцу двух и более быстро следующих друг за другом раздражений.

Рис. 25. Суммация сокращений икроножной мышцы лягушки: 1 – кривая одиночного сокращения в ответ на первое раздражение расслабленной мышцы; 2 – кривая одиносного сокращения той же мышцы в ответ на второе раздражние; 3 – кривая суммированного сокращения, полученного в результате спаренного раздражения сокращающейся мышцы (обозначено стрелками )
При этом раздражения должны поступать в период предыдущего сокращения. Увеличение амплитуды сокращений объясняется увеличением концентрации Са 2+ в гиалоплазме при повторном возбуждении мышечных волокон, поскольку Са-помпа не успевает возвращать его в СПР. Са 2+ обеспечивает увеличение числа зон зацепления миозиновых мостиков с нитями актина.
Если повторные импульсы или раздражения поступают в фазу расслабления мышц, возникает зубчатый тетанус . Если повторные раздражения приходятся на фазу укорочения, возникает гладкий тетанус (рис. 26).

Рис. 26. Сокращение икроножной мышцы лягушки при различной частоте раздражения седалищного нерва: 1 – одиночное сокращение (частота 1 Гц); 2,3 – зубчатый тетанус (15-20 Гц); 4,5 – гладкий тетанус (25-60 Гц); 6 – расслабление при пессимальной частоте раздражения (120 Гц)
Амплитуда сокращения и величина напряжения, развиваемые мышечными волокнами при гладком тетанусе, обычно в 2-4 раза больше, чем при одиночном сокращении. Тетаническое сокращение мышечных волокон, в отличие от одиночных сокращений, быстрее вызывает их утомление.
При возрастании частоты стимуляции нерва или мышцы амплитуда гладкого тетануса увеличивается. Максимальный тетанус получил название оптимума. Увеличение тетануса объясняется накоплением Са 2+ в гиалоплазме. При дальнейшем увеличении частоты стимуляции нерва (около 100 Гц) мышца расслабляется вследствие развития блока проведения возбуждения в нервно-мышечных синапсах - пессимум Введенского (частота раздражения пессимальная ) (см. рис. 26). Пессимум Введенского можно получить и при прямом, но более частом раздражении мышцы (около 200 имп./с), однако при этом для чистоты эксперимента следует заблокировать нервно-мышечные синапсы. Если после возникновения пессимума уменьшить частоту стимуляции до оптимальной, то амплитуда мышечного сокращения мгновенно возрастает - свидетельство того, что пессимум не является результатом утомления мышцы или истощением энергетических ресурсов.
Скелетные мышцы в условиях опыта отвечают на одиночное раздражение одиночным сокращением. Однако в целом организме одиночные сокращения свойственны только сердечной мышце, которая сокращается в ответ на одиночные импульсы, поступающие к ней из синусного узла.
В зависимости от частоты импульсации мышца сокращается по разному:
Для того чтобы понять механизм тетануса, необходимо изучить одиночное мышечное сокращение, которое является непременной составной единицей каждого их них.
Изучение одиночного мышечного сокращения можно провести, если записать его в развернутом виде, используя для этого быстро вращающийся кимограф.
В естественных условиях мышцы сокращаются под влиянием ритмических импульсов, получаемых из ЦНС. Импульсы следуют с частотой большей, чем период одиночного мышечного сокращения, т.е. мышца, не успев расслабиться, получает следующий. В мышцах возникает явление суммации, в результате которого они приходят в состояние длительного укорочения, называемого тетанусом . Экспериментально тетанус можно получить на икроножной мышце лягушки при воздействии ритмического раздражителя. При частоте, когда каждый последующий раздражитель попадает в фазу расслабления мышцы, получается зубчатый тетанус .
При частоте раздражения, когда каждый последующий импульс попадает в фазу укорочения мышцы, возникает длительное непрерывное сокращение, которое называется гладким тетанусом.
АВ - латентный период - 0,01 сек.
ВС - период укорочения - 0,04 сек.
СД - период расслабления - 0,05 сек.
АД - период сокращения мышцы - 0,1 сек.
При возбуждении и сокращении мышцы изменяется ее возбудимость.
Как только на мышцы подействовал пороговый раздражитель, в мышце возникло возбуждение, а возбудимость ее пропала, это будет абсолютная рефрактерная фаза, т. е. абсолютная невозбудимость и если в этот момент наносить дополнительные раздражения мышцы на них не будут отвечать, эта фаза длиться 0,001 - 0,003 секунды. Затем возбудимость постепенно восстанавливается и на новые дополнительные более сильные раздражения мышцы отвечает слабым сокращением. Это относительно рефрактерная фаза, длиться она 0,009 - 0,007 секунды. Обе эти фазы укладываются в латентный период. После относительно рефрактерной фазы возбудимость в мышце не только восстанавливается, но и становиться значительно выше исходной - экзальтационная фаза - 0,018 секунды. Затем возбудимость возвращается к исходной величине.
Различают следующие режимы мышечных сокращений:
Изотоническое - сокращение, при котором происходит укорочение мышечных волокон, но их напряжение не меняется.
Изометрическое - сокращение, при котором длина волокон не уменьшается, но их напряжение возрастает.
Ауксотоническое - сокращение, при котором изменяются и длина и напряжение мышц. Такой режим сокращения характерен для работающих мышц в целом организме. Первые два можно получить только в эксперименте.
рис. 2.4. Электрическое раздражение и мышечный ответ. Сверху показаны электрические импульсы, снизу - ответ мышцы
Если стимулировать коротким электрическим импульсом, спустя небольшой латентный период происходит ее . Такое сокращение называется «одиночное сокращение мышцы». Одиночное мышечное сокращение длится около 10-50 мс, причем оно достигает максимальной силы через 5-30 мс.
Каждое отдельное мышечное волокно подчиняется закону «все или ничего», т. е. при силе раздражения выше порогового уровня происходит полное сокращение с максимальной для данного волокна силой, а ступенчатое повышение силы сокращения по мере увеличения силы раздражения невозможно. Поскольку смешанная мышца состоит из множества волокон с различным уровнем чувствительности к возбуждению, сокращение всей мышцы может быть ступенчатым в зависимости от силы раздражения, при этом при сильных раздражениях происходит активация глубжележащих мышечных волокон.
Суперпозиция и тетанус
Однократное электрическое раздражение (рис. 2.4, вверху) ведет к единичному мышечному сокращению (рис. 2.4, внизу). Два близко друг за другом следующих раздражения накладываются друг на друга (это называется «суперпозиция», или суммация сокращений), что ведет к более сильному мышечному ответу, близкому к максимальному. Серия часто повторяющихся электрических раздражений вызывает возрастающие по силе мышечные сокращения, в результате чего не происходит должного расслабления мышцы. Если частота электрических импульсов выше частоты слияния, то единичные раздражения сливаются в одно и вызывают тетанус мышцы (тетаническое сокращение) - устойчивое достаточно длительное напряжение сокращенной мышцы.
Формы сокращений

Рис. 2.5. Формы мышечных сокращений. Слева схематически представлено укорочение саркомеров, в середине - изменения силы и длины, справа - пример сокращений
Выделяют различные функциональные формы мышечных сокращений (рис. 2.5).
- При изотоническом сокращении мышца укорачивается, однако ее внутреннее напряжение (тонус!) остается неизменным во всех фазах рабочего цикла. Типичным примером изотонического мышечного сокращения является динамическая мышечная работа сгибателей и разгибателей без существенных изменений внутримышечного напряжения, например подтягивание.
- При изометрическом сокращении мышечная длина не изменяется, а сила мышцы проявляется в повышении ее напряжения. Типичным примером изометрического сокращения является статическая мышечная активность при поднимании тяжестей (удерживание штанги).
- Чаще всего наблюдаются комбинированные варианты сокращения мышц. Например, комбинированное сокращение, при котором мышцы сначала сокращаются изометрически, а затем изотонически, как при поднятии тяжести, называют удерживающим сокращением .
- Установочным (изготовочным) называют сокращение, при котором, наоборот, после начального изотонического сокращения следует изометрическое. Примером является ротационное движение руки с рычагом - затягивание винта с помощью гаечного ключа или отвертки.
- Различные формы мышечных сокращений выделяют для их описания и систематизации. На самом деле в большинстве динамических спортивных движений происходит как укорочение мышцы, так и повышение напряжения (тонуса) мышц - ауксотонические сокращения .
Использованные здесь термины нетипичны для русской литературы по мышечной активности. В отечественной литературе принято выделять следующие типы сокращений.
- Концентрическое сокращение - вызывающее укорачивание мышцы и перемещение места прикрепления ее к кости, при этом движение конечности, обеспечиваемое сокращением данной мышцы, направлено против преодолеваемого сопротивления, например силы тяжести.
- Эксцентрическое сокращение - возникает при удлинении мышцы во время регулирования скорости движения, вызванного другой силой, или в ситуации, когда максимального усилия мышцы не хватает для преодоления противодействующей силы. В результате движение происходит в направлении воздействия внешней силы.
- Изометрическое сокращение - усилие, противодействующее внешней силе, при котором длина мышцы не изменяется и движения в суставе не происходит.
- Изокинетическое сокращение - сокращение мышцы с одинаковой скоростью.
- Баллистическое движение - быстрое движение, включающее: а) концентрическое движение мышц-агонистов в начале движения; б) инерционное движение во время минимальной активности; в) эксцентрическое сокращение для замедления движения.
Механизм скольжения филаментов

рис. 2.6 Схема образования поперечных связей - молекулярной основы сокращения саркомера
Укорочение мышцы происходит за счет укорочения образующих ее саркомеров, которые, в свою очередь, укорачиваются за счет скольжения относительно друг друга актиновых и миозиновых филаментов (а не укорочения самих белков). Теория скольжения филаментов была предложена учеными Huxley и Hanson (Huxley, 1974; рис. 2.6). (В 1954 г. две группы исследователей - X. Хаксли с Дж. Хэнсон и А. Хаксли с Р. Нидергерке - сформулировали теорию, объясняющую мышечное сокращение скольжением нитей. Независимо друг от друга они обнаружили, что длина диска А оставалась постоянной в расслабленном и укороченном саркомере. Это позволило предположить, что есть два набора нитей - актиновые и миозиновые, причем одни входят в промежутки между другими, и при изменении длины саркомера эти нити каким-то образом скользят друг по другу. Сейчас эта гипотеза принята почти всеми.)
Актин и миозин - два сократительных белка, которые способны вступать в химическое взаимодействие, приводящее к изменению их взаимного расположения в мышечной клетке. При этом цепочка миозина прикрепляется к актиновой нити с помощью целого ряда особых «головок», каждая из которых сидит на длинной пружинистой «шее». Когда происходит сцепление между миозиновой головкой и актиновой нитью, конформация комплекса этих двух белков изменяется, миозиновые цепочки продвигаются между актиновыми нитями и мышца в целом укорачивается (сокращается). Однако, чтобы химическая связь между головкой миозина и активной нитью образовалась, необходимо подготовить этот процесс, поскольку в спокойном (расслабленном) состоянии мышцы активные зоны белка актина заняты другим белком - тропохмиозином, который не позволяет актину вступить во взаимодействие с миозином. Именно для того, чтобы убрать тропомиозиновый «чехол» с актиновой нити, требуется быстрое выливание ионов кальция из цистерн саркоплазматического ретикулума, что происходит в результате прохождения через мембрану мышечной клетки потенциала действия. Кальций изменяет конформацию молекулы тро-помиозина, в результате чего активные зоны молекулы актина открываются для присоединения головок миозина. Само это присоединение осуществляется с помощью так называемых водородных мостиков, которые очень прочно связывают две белковые молекулы - актин и миозин - и способны в таком связанном виде находиться очень долго.
Для отсоединения миозиновой головки от актина необходимо затратить энергию аденозинтрифосфа-та (АТФ), при этом миозин выступает в роли АТФазы (фермента, расщепляющего АТФ). Расщепление АТФ на аденозиндифосфат (АДФ) и неорганический фосфат (Ф) высвобождает энергию, разрушает связь между актином и миозином и возвращает головку миозина в исходное положение. В дальнейшем между актином и миозином могут снова образовываться поперечные связи.
При отсутствии АТФ актин-миозиновые связи не разрушаются. Это и является причиной трупного окоченения (rigor mortis) после смерти, т. к. останавливается выработка АТФ в организме - АТФ предотвращает мышечную ригидность.
Даже при мышечных сокращениях без видимого укорочения (изометрические сокращения, см. выше) активируется цикл формирования поперечных связей, мышца потребляет АТФ и выделяет тепло. Головка миозина многократно присоединяется на одно и то же место связывания актина, и вся система миофиламентов остается неподвижной.
Внимание : Сократительные элементы мышц актин и миозин сами по себе не способны к укорочению. Мышечное укорочение является следствием взаимного скольжения миофиламентов относительно друг друга (механизм скольжения филаментов).
Как же образование поперечных связей (водородных мостиков) переходит в движение? Одиночный саркомер за один цикл укорачивается приблизительно на 5-10 нм, т.е. примерно на 1 % своей общей длины. За счет быстрого повторения цикла поперечных связей возможно укорочение на 0,4 мкм, или 20% своей длины. Поскольку каждая миофибрилла состоит из множества саркомеров и во всех них одновременно (но не синхронно) образуются поперечные связи, суммарно их работа приводит к видимому укорочению всей мышцы. Передача силы этого укорочения происходит через Z-линии миофибрилл, а также концы сухожилий, прикрепленных к костям, в результате чего и возникает движение в суставах, через которые мышцы реализуют перемещение в пространстве частей тела или продвижение всего тела.
Связь между длиной саркомера и силой мышечных сокращений

Рис. 2.7. Зависимость силы сокращений от длины саркомера
Наибольшую силу сокращений мышечные волокна развивают при длине 2-2,2 мкм. При сильном растяжении или укорочении саркомеров сила сокращений снижается (рис. 2.7). Эту зависимость можно объяснить механизмом скольжения филаментов: при указанной длине саркомеров наложение миозиновых и актиновых волокон оптимально; при большем укорочении миофиламенты перекрываются слишком сильно, а при растяжении наложение миофиламентов недостаточно для развития достаточной силы сокращений.

рис. 2.9 Влияние предварительного растяжения на силу сокращения мышцы. Предварительное растяжение повышает напряжение мышцы. Результирующая кривая, описывающая взаимоотношения длины мышцы и силы ее сокращения при воздействии активного и пассивного растяжения, демонстрирует более высокое изометрическое напряжение, чем в покое
Важным фактором, влияющим на силу сокращений, является величина растяжения мышцы. Тяга за конец мышцы и натяжение мышечных волокон называются пассивным растяжением. Мышца обладает эластическими свойствами, однако в отличие от стальной пружины зависимость напряжения от растяжения не линейна, а образует дугообразную кривую. С увеличением растяжения повышается и напряжение мышцы, но до определенного максимума. Кривая, описывающая эти взаимоотношения, называется кривой растяжения в покое .
Данный физиологический механизм объясняется эластическими элементами мышцы - эластичностью сарколеммы и соединительной ткани, располагающимися параллельно сократительным мышечным волокнам.
Также при растяжении изменяется и наложение друг на друга миофиламентов, однако это не оказывает влияния на кривую растяжения, т. к. в покое не образуются поперечные связи между актином и миозином. Предварительное растяжение (пассивное растяжение) суммируется с силой изометрических сокращений (активная сила сокращений).